Page 1 of 33
OP4.1 | Corneal Anatomy, Physiology and Transparency — SDL Guide
Learning Objectives
- Describe the five-layered structure of the cornea and the ultrastructure of each layer
- Explain the three principal mechanisms responsible for corneal transparency
- Describe the metabolic physiology of the cornea including oxygen supply, glucose metabolism, and innervation
- Correlate physiological failure states with clinical presentations such as corneal oedema and neurotrophic keratitis
- Outline the clinical tools used to assess corneal structure and endothelial function
INSTRUCTIONS
The cornea is the principal refracting surface of the eye, contributing approximately 43 dioptres of the eye's total ~60-dioptre optical power. Its transparency — essential for vision — is not a passive property but a physiologically maintained state that depends on precise ultrastructural organisation and active cellular pumping. Understanding corneal anatomy and physiology is foundational to every clinical ophthalmology decision: from reading a slit-lamp correctly, to understanding why a corneal ulcer threatens vision, to knowing why extended contact lens wear can be dangerous.
References
- Khurana AK. Comprehensive Ophthalmology, 7th ed. Ch 5: Diseases of the Cornea (textbook)
- Parsons' Diseases of the Eye, 23rd ed. Ch 9: The Cornea (textbook)
Version 2.0 | NMC CBUC 2024
CLINICAL SCENARIO
A 45-year-old long-distance truck driver presents with a painless, gradually progressive haziness in both eyes over 18 months. His visual acuity is 6/60 in the right eye and 6/36 in the left. Slit-lamp examination reveals diffuse greyish corneal stromal oedema with epithelial microcysts. His specular microscopy shows an endothelial cell count of 480 cells/mm². The corneas look 'foggy' from within. His consultant says: 'The pump has failed.' What does that mean — and what is the pump, where does it live, and what exactly is it pumping?
WHY THIS MATTERS
Every corneal disease you will encounter in clinical practice — keratitis, trauma, dystrophy, post-surgical complications — can ultimately be traced to disruption of the anatomical layers or the physiological mechanisms described in this module. The clarity of the cornea is not structural luck; it is a biological achievement that your patients' endothelial cells perform moment-to-moment. When you examine a cornea with a slit-lamp and describe what you see ('epithelial irregularity', 'stromal haze', 'Descemet's folds'), you are reading the language of anatomy. When you counsel a patient about soft contact lens wear duration, you are applying corneal physiology. This module builds the anatomical and physiological vocabulary that all subsequent ophthalmology topics depend on.
RECALL
Before entering corneal layers, bring forward your anatomy and histology: the cornea is continuous with the sclera at the corneoscleral limbus. From embryology, recall that the corneal epithelium derives from surface ectoderm, while the stroma and endothelium are derived from neural crest cells that migrate in during the 5th-6th week of embryogenesis. From physiology, remember that avascular tissues receive nutrients by diffusion — oxygen dissolving into the tear film from the atmosphere, and glucose diffusing from the aqueous humour. From optics, the cornea provides approximately two-thirds of the total refracting power of the eye because of the large refractive index change at its air-tear interface.
Corneal Layers and Their Ultrastructure
The cornea is composed of five distinct layers, each with a unique cellular composition and functional role. Together they form a transparent, dome-shaped, avascular tissue approximately 11.5–12 mm in horizontal diameter and 520–540 µm thick at the centre (slightly thicker at the periphery). Understanding each layer individually is essential, because different diseases target different layers and the clinical signs on slit-lamp examination reflect the layer involved.
The outermost layer, the corneal epithelium, is a non-keratinised, stratified squamous epithelium consisting of 5–6 cell layers. It comprises three cell types: superficial flattened squamous cells (with microplicae and microvilli that stabilise the tear film), wing cells in the middle tiers, and a single layer of columnar basal cells anchored to the underlying basement membrane via hemidesmosomes. Basal cells are mitotically active; new cells are generated here and migrate superficially, desquamating into the tear film every 7–10 days. The entire epithelium is renewed approximately every week. Crucially, stem cells at the limbus (the palisades of Vogt) replenish the basal cell population; limbal stem cell deficiency is a significant clinical entity causing conjunctivalisation of the corneal surface.
Immediately beneath the epithelium lies Bowman's layer (also called the anterior limiting lamina), an acellular condensation of randomly arranged collagen fibrils (Types I and V) approximately 8–14 µm thick. It has no regenerative capacity — once destroyed by trauma or infection, it heals with an opaque fibrous scar. Its exact function remains debated, but it appears to provide mechanical resistance and a barrier to epithelial ingrowth.
The stroma constitutes approximately 90% of the total corneal thickness (~470–500 µm of the 520 µm total). It is composed of about 200 lamellae of Type I collagen fibrils arranged in an orthogonal lattice with extraordinary regularity. Interspersed within the lamellae are keratocytes (corneal fibroblasts), which synthesise collagen and proteoglycans. The ground substance is rich in keratan sulphate and dermatan sulphate proteoglycans, which space the collagen fibrils precisely and contribute to transparency. The stroma is the layer most affected by bacterial and fungal ulcers, where inflammatory infiltration and melting produce the characteristic clinical signs.
Descemet's membrane is the thick basement membrane secreted by the endothelium, typically 10–12 µm thick in adults. It has an anterior banded zone (laid down in utero) and a posterior non-banded zone (added throughout life). It is remarkably resistant to collagenases and persists in the stroma even when the overlying layers are destroyed — a fact exploited in deep anterior lamellar keratoplasty (DALK), where the surgeon removes epithelium and stroma down to Descemet's membrane. Haab's striae are horizontal breaks in Descemet's membrane seen in congenital glaucoma due to stretching of the enlarged cornea.
The innermost layer is the corneal endothelium — a single layer of hexagonal cells (approximately 2,500–3,000 cells/mm² in young adults) resting on Descemet's membrane. These cells are almost entirely post-mitotic; they cannot regenerate in vivo after the first year of life. When endothelial cells die, the neighbours enlarge and spread to cover the defect (pleomorphism and polymegethism). Below a critical cell density of approximately 500 cells/mm², the pump fails and corneal decompensation occurs. This irreplaceable nature of the endothelium explains why endothelial cell counting by specular microscopy is so important before intraocular surgery.
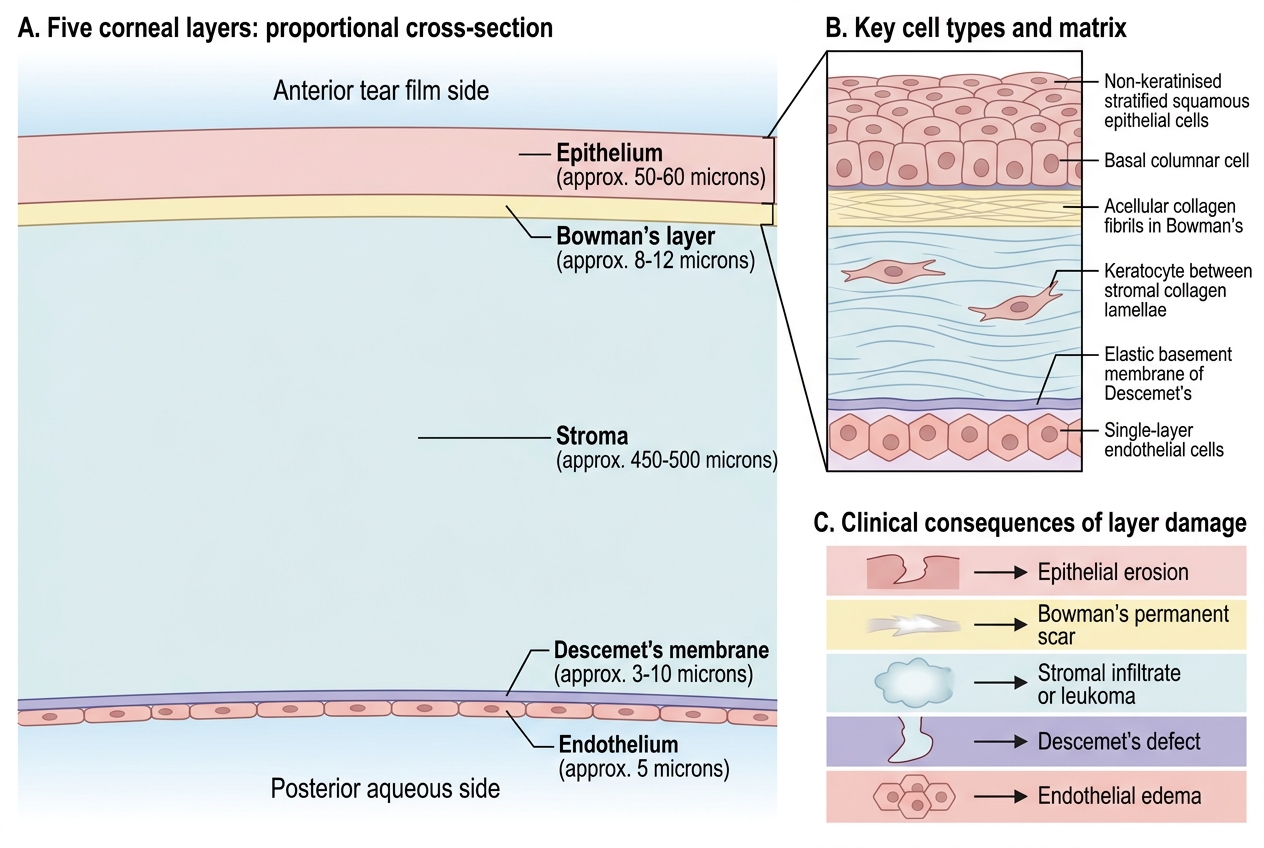
Five Layers of the Cornea
| Layer | Thickness | Key Cell Type | Primary Function | Consequence of Damage |
|---|---|---|---|---|
| Epithelium | 50–60 µm | Non-keratinised squamous; basal columnar cells | Barrier; tear film stabilisation | Erosion, recurrent erosion syndrome |
| Bowman's layer | 8–14 µm | Acellular (collagen fibrils) | Mechanical resistance | Permanent scar if destroyed |
| Stroma | ~470–500 µm | Keratocytes in collagen lamellae | Structural support; transparency | Infiltrate, ulcer, scar (leukoma) |
| Descemet's membrane | 10–12 µm | Acellular (basement membrane) | Endothelial substrate | Haab's striae; DM detachment post-DALK |
| Endothelium | Single cell layer | Post-mitotic hexagonal cells | Active dehydration (pump) | Corneal oedema → bullous keratopathy |
SELF-CHECK
A patient undergoes penetrating eye trauma that destroys the Bowman's layer. What is the likely long-term visual consequence?
A. Full recovery without scarring because Bowman's layer regenerates from keratocytes
B. Permanent corneal scar at that site because Bowman's layer has no regenerative capacity
C. Corneal oedema because Bowman's layer pumps fluid from the stroma
D. Recurrent corneal erosion because Bowman's layer anchors the basal epithelium
Reveal Answer
Answer: B. Permanent corneal scar at that site because Bowman's layer has no regenerative capacity
Bowman's layer is acellular and cannot regenerate once damaged. Destruction results in fibrous scarring visible as a permanent corneal opacity. The pumping function belongs to the endothelium, not Bowman's layer. Recurrent erosion relates to defects in epithelial-basement membrane adhesion.
Factors Maintaining Corneal Transparency
Corneal transparency is one of the most precisely maintained physiological states in the body. Despite being a living tissue composed of cells, collagen, and proteoglycans, the cornea transmits 90% of visible light with negligible scatter. Three interdependent mechanisms are responsible: avascularity, regular collagen spacing, and active dehydration by the endothelial pump.
Avascularity is the first and most obvious requirement. Blood vessels are opaque because red blood cells absorb light, and vascular walls scatter light. The cornea is kept avascular by a careful balance between angiogenic and anti-angiogenic factors at the limbus. VEGF (vascular endothelial growth factor) is produced by corneal cells under hypoxic stress (such as extended contact lens wear), and the cornea normally counteracts this with anti-VEGF factors including PEDF (pigment epithelium-derived factor) and sFLT1. When this balance is disrupted — by chronic inflammation, infection, or prolonged hypoxia — corneal neovascularisation develops, carrying blood vessels and, with them, opacity into the tissue. This is a critical reason why microbial keratitis must be treated aggressively.
Regular collagen fibril spacing is the structural basis of transparency, elegantly explained by the Maurice lattice theory (1957). In the corneal stroma, Type I collagen fibrils are arranged with a remarkably uniform diameter (~24 nm) and spacing (~60 nm centre-to-centre), maintained by keratan sulphate proteoglycans. This spacing is smaller than half the wavelength of visible light (400–700 nm). As a result, light scattered by each fibril is out of phase with scattered light from neighbouring fibrils — they undergo destructive interference, cancelling the scattered component. Only the transmitted beam passes forward undisturbed. This is directly analogous to how anti-reflection coatings work in optics. When the stroma is oedematous (as in Fuchs' dystrophy or bullous keratopathy), water disrupts this regular spacing, scattered light adds constructively, and the stroma becomes hazy.
⚑ AI image — pending faculty review (auto-QA score 5/10; best of 3 attempts)
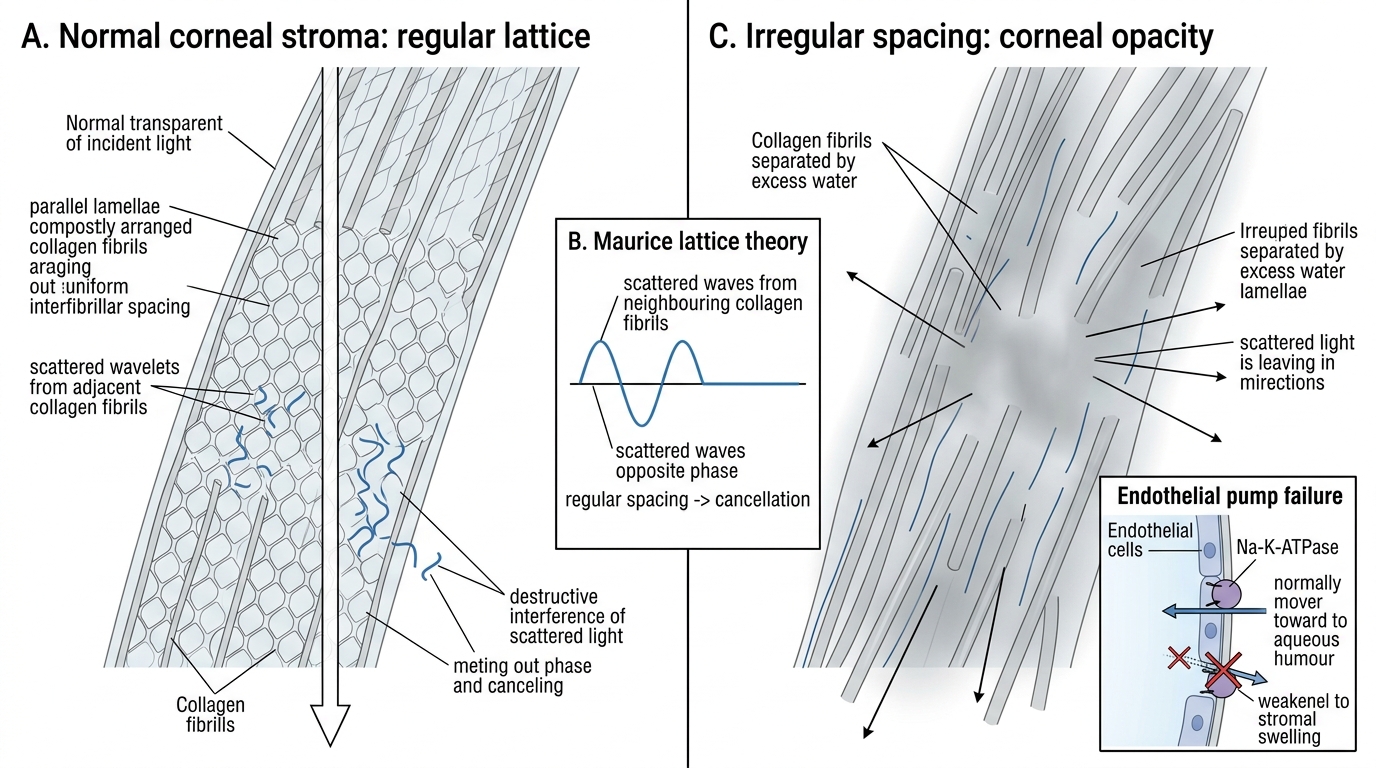
Maurice Lattice Theory of Corneal Transparency
Active dehydration by the endothelial pump is the third and clinically most important mechanism. The stroma has a natural tendency to swell (imbibition) because its proteoglycans are hydrophilic. The endothelium combats this by continuously pumping water out of the stroma into the aqueous humour. This pump is powered by Na-K-ATPase on the lateral cell membranes and carbonic anhydrase (which generates bicarbonate ions that create an osmotic gradient). The normal water content of the stroma is approximately 78%; if the pump fails, this rises and the stroma swells and scatters light. The epithelium provides an additional barrier against tears entering the stroma from the front. The combined effect of the epithelial barrier and the endothelial pump is sometimes called the pump-leak model of corneal deturgescence. When endothelial cell density falls below ~500 cells/mm², decompensation occurs — the pump cannot keep up with the leak, and the cornea becomes progressively oedematous.
SELF-CHECK
According to Maurice's lattice theory, why does regular collagen fibril spacing maintain transparency?
A. The regular spacing reflects light forward in a focused beam, increasing transmission
B. Collagen fibrils are too thin to scatter visible light at 24 nm diameter
C. Light scattered by each fibril is out of phase with scatter from neighbours, causing destructive interference that cancels scattered light
D. Regular spacing prevents leukocyte infiltration, maintaining the avascular state
Reveal Answer
Answer: C. Light scattered by each fibril is out of phase with scatter from neighbours, causing destructive interference that cancels scattered light
Maurice's lattice theory states that the regular spacing of collagen fibrils (< half-wavelength of visible light) means scattered light waves from adjacent fibrils are out of phase and cancel by destructive interference. The diameter of individual fibrils is indeed small, but it is the spacing regularity — not small size alone — that creates transparency. When oedema disrupts the regular spacing, the interference is no longer perfectly destructive and the stroma becomes hazy.
Corneal Physiology: Metabolism, Oxygen and Nutrition
Being avascular, the cornea has evolved special routes to obtain oxygen, nutrients, and neural signals. Understanding these routes is not merely academic — it directly explains why extended contact lens wear is harmful, why diabetic patients develop corneal complications, and how the corneal reflex works.
Oxygen supply is the most critical physiological need, particularly for the metabolically active epithelial cells. The cornea obtains oxygen primarily from the atmosphere, dissolved in the pre-corneal tear film. In the open eye, the partial pressure of oxygen in tears (approximately 155 mmHg at the corneal surface) is sufficient to sustain aerobic metabolism. During sleep — when the eyelids are closed — the conjunctival circulation provides oxygen at much lower partial pressure (~55 mmHg), a condition the cornea tolerates with a shift towards anaerobic glycolysis and mild lactic acid accumulation. Contact lenses act as a barrier to atmospheric oxygen; soft contact lenses with low oxygen transmissibility (Dk/t) create hypoxic conditions at the epithelium, inducing VEGF and risking neovascularisation. Modern silicone hydrogel lenses have much higher Dk/t values and are safer for extended wear.
Glucose and nutrients are supplied primarily from the aqueous humour (for the endothelium and posterior stroma), supplemented by the limbal blood vessels at the periphery (for the peripheral stroma and limbal epithelium). The epithelium metabolises glucose mainly via aerobic glycolysis; under hypoxia it shifts to anaerobic glycolysis, producing lactate that can cause stromal acidosis and epithelial oedema. The lens and cornea compete for glucose from the aqueous; in poorly controlled diabetes, fluctuating glucose levels affect both.
Sensory innervation is provided by the long ciliary nerves, branches of the nasociliary nerve (ophthalmic division, V1 of the trigeminal nerve). The cornea is, per unit area, the most densely innervated tissue in the human body, with a nerve density approximately 300–400 times that of skin. This dense innervation underlies the extreme sensitivity of the corneal reflex — the afferent limb is V1, the efferent is the facial nerve (CN VII) to orbicularis oculi. Clinically, the corneal sensitivity test (using a wisp of cotton wool or Cochet-Bonnet aesthesiometer) is used to detect early peripheral neuropathy, to diagnose herpes simplex keratitis (which reduces corneal sensation), and to monitor neurotrophic keratitis — a sight-threatening condition in which loss of trophic signals from nerve fibres leads to poor epithelial healing, recurrent erosions, and eventually sterile corneal ulceration.