Page 1 of 25
PA14.1 | Iron Metabolism: Absorption, Transport, Storage, Regulation — SDL Guide
Learning Objectives
- Describe the distribution of total body iron across the three pools — functional (haemoglobin, myoglobin, enzymes), storage (ferritin, hemosiderin), and transport (transferrin) — and explain why these pools differ in clinical detectability
- Trace the pathway of dietary iron from duodenal lumen to erythroblast: DMT1 → enterocyte → ferroportin → plasma → transferrin → TfR-mediated endocytosis
- Explain transferrin saturation and TIBC as derived measures and interpret them in a given iron-study panel
- Distinguish ferritin from hemosiderin as storage forms and explain why serum ferritin is the best single marker of iron stores — and when it becomes unreliable
- Describe hepcidin's role as the master regulator of systemic iron homeostasis and predict the direction of hepcidin change in iron deficiency versus chronic inflammation
- Sequence the three stages of progressive iron depletion and identify which laboratory parameter falls first in each stage
INSTRUCTIONS
You already classified anaemia by MCV in the previous cluster. You know microcytic anaemia exists. Now the question is: why does iron deficiency produce that pattern? The answer lives in iron metabolism — a beautifully regulated system that you studied in Year-1 Biochemistry, but in fragments. This module re-assembles those fragments into one coherent picture. By the time you finish, when you see 'low Hb, low MCV, low ferritin' on a CBC report, you will know exactly which pool is depleted, why it depleted, and what the body tried to do about it. That understanding is the foundation for every microcytic anaemia you will diagnose for the rest of your career.
References
- Robbins and Cotran Pathologic Basis of Disease, 10th ed., Chapter 14 — Red Blood Cell Disorders (textbook)
- Hoffbrand's Essential Haematology, 8th ed., Chapter 2 — Iron Metabolism and Clinical Aspects of Iron Deficiency (textbook)
- WHO/IAEA Serum Ferritin Concentrations for the Assessment of Iron Status and Iron Deficiency in Populations (2020) (guideline)
- Muckenthaler MU et al. A Red Carpet for Iron Metabolism. Cell. 2017;168(3):344-361. (review)
Version 2.0 | NMC CBUC 2024
CLINICAL SCENARIO
A 28-year-old woman in her second trimester presents with progressive fatigue and breathlessness on climbing stairs. Her CBC: Hb 8.2 g/dL, MCV 68 fL, MCHC 28 g/dL. You order an iron study panel and get back: serum iron 40 µg/dL (low), TIBC 480 µg/dL (high), transferrin saturation 8% (low), serum ferritin 4 ng/mL (very low). You know this is iron deficiency anaemia. But do you know why the body ran out of iron while doing everything physiologically possible to conserve it? The answer requires understanding a system that processes 25 mg of iron per day with extraordinary precision — and gets only 1-2 mg from food.
WHY THIS MATTERS
Iron deficiency anaemia is the world's most prevalent nutritional deficiency, affecting over 1.2 billion people globally. In India, more than 50% of pregnant women and 60% of under-5 children are anaemic — the majority from iron deficiency. As a clinician, you will order iron studies in almost every medicine, obstetrics, and paediatrics posting. The 4-parameter iron panel (serum iron, TIBC, transferrin saturation, serum ferritin) is one of the most commonly misinterpreted sets of results in clinical practice. Knowing the physiology turns interpretation from memory into reasoning. It also sets up the entire differential diagnosis for microcytic anaemia — because each different cause (IDA, anaemia of chronic disease, thalassaemia trait, sideroblastic anaemia) perturbs the system at a different point.
RECALL
From Year-1 Biochemistry (PA/BI cross-reference): You covered iron as a cofactor for haemoglobin synthesis — specifically the role of protoporphyrin IX + ferrous iron (Fe²⁺) forming haem, and haem + globin chains forming haemoglobin. You may also recall ferritin as the storage form and transferrin as the serum transport protein. In Year-1 Physiology, you covered erythropoiesis, the role of erythropoietin (EPO), and reticulocyte production. If these feel distant, take 2 minutes to recall: (a) haem synthesis occurs in the mitochondria and cytoplasm of erythroblasts, (b) iron enters the protoporphyrin ring at the final step via ferrochelatase, and (c) without iron, that final step stalls — haemoglobin cannot be made. Today's module answers: where does that iron come from, how does it get to the erythroblast, and how does the body know how much to absorb?
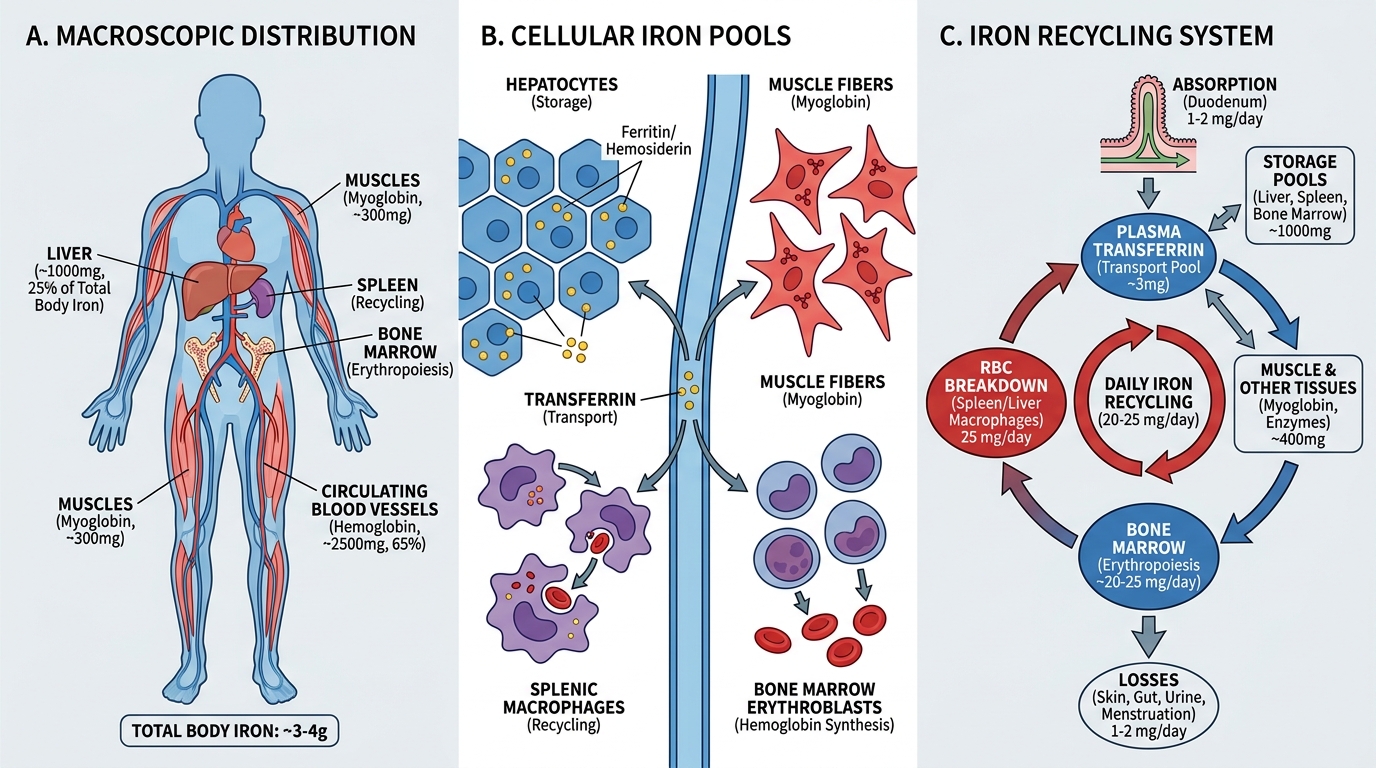
Iron Distribution: Three Pools, One System
The average adult contains 3–4 g of total body iron, distributed across three functional pools:
1. Functional pool (~65–70% of total)
- Haemoglobin iron: ~2,500 mg in circulating RBCs. Each haemoglobin molecule carries four haem groups; each haem contains one Fe²⁺ ion. This is why haemoglobin mass is the largest iron consumer in the body.
- Myoglobin: ~300 mg in muscle. Binds oxygen for aerobic metabolism.
- Haem-containing enzymes: ~8 mg — cytochromes, catalase, peroxidase.
2. Storage pool (~20–25% of total)
- Ferritin: ~800–1200 mg in hepatocytes, macrophages (reticuloendothelial system), and enterocytes. Iron is stored here in a soluble, mobilisable form within the apoferritin shell.
- Hemosiderin: ~200–300 mg — an insoluble aggregate of degraded ferritin + iron. Visible as golden-brown granules on H&E; confirmed by Perls' Prussian Blue staining (a critical histology skill you will use in bone marrow biopsies).
3. Transport pool (<0.1% of total — but the clinically measured fraction)
- Transferrin-bound iron: ~3–4 mg at any time. This tiny pool is in constant flux, delivering iron from absorption sites and macrophages to erythropoietic marrow.
The critical insight: 25 mg of iron is recycled daily from senescent RBC destruction by splenic macrophages (reticuloendothelial system). Dietary absorption adds only 1–2 mg/day — barely replacing insensible losses (sweat, desquamation, menstruation). This means the body is almost entirely dependent on efficient iron recycling. When recycling fails — or losses exceed recycling — the storage pool depletes first.
Iron Distribution: Three Pools, One System
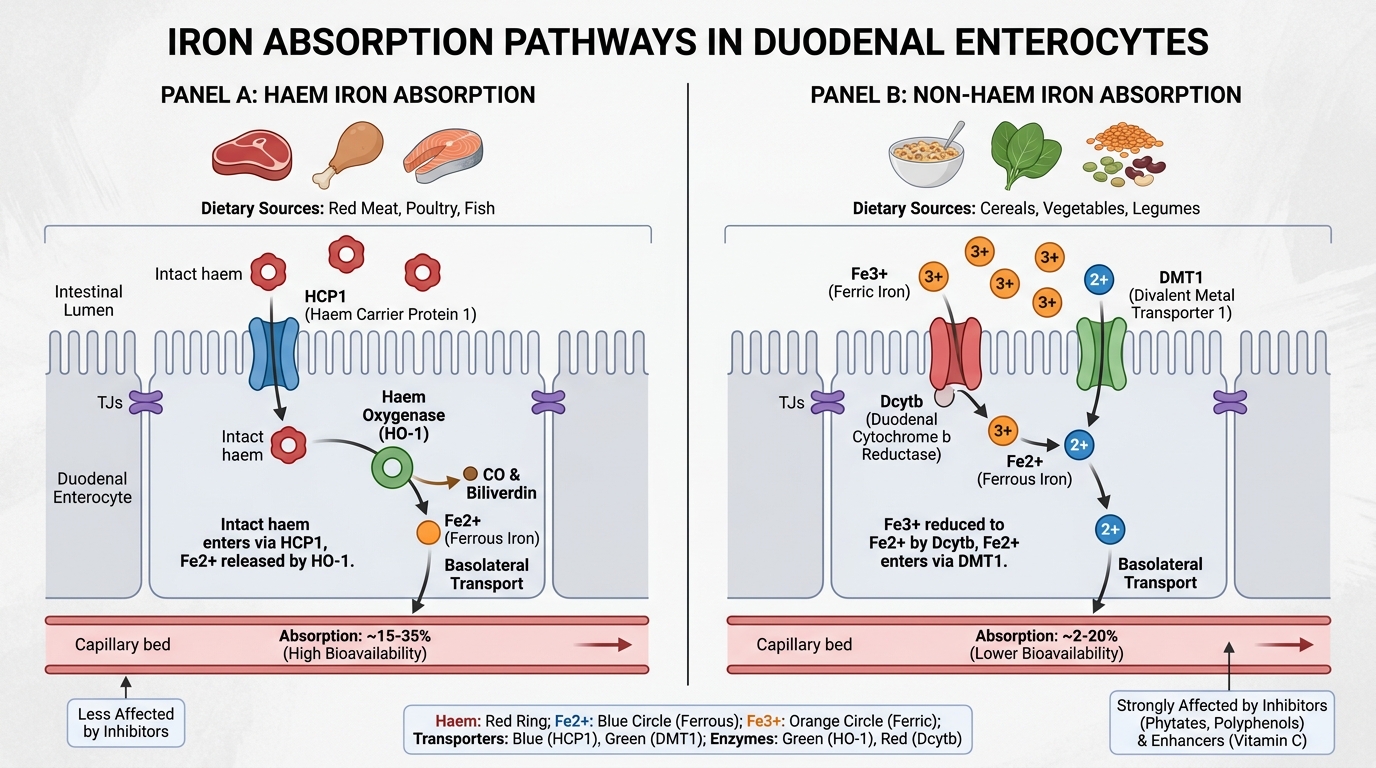
Iron Absorption: The Duodenal Gateway
The duodenum and proximal jejunum are the exclusive sites of regulated iron absorption. The body cannot excrete iron — the only way iron leaves is through blood loss, desquamation, or menstruation. Therefore, the entry gate at the duodenum is the primary regulatory point.
Two dietary iron forms, two pathways:
| Feature | Haem iron | Non-haem iron |
|---|---|---|
| Sources | Red meat, poultry, fish | Cereals, legumes, vegetables, dairy |
| % of diet | 10–15% | 85–90% |
| Absorption rate | 20–30% | 1–8% |
| Affected by enhancers/inhibitors | Minimally | Significantly |
| Absorption mechanism | Haem taken intact into enterocyte → HCP1 transporter; haem oxygenase releases Fe²⁺ inside | Requires reduction Fe³⁺ → Fe²⁺ by Dcytb; then DMT1 transport |
The non-haem pathway in detail (the one that matters most for iron deficiency):
1. Gastric acid reduces luminal pH → Fe³⁺ solubilised → duodenal cytochrome b (Dcytb, a brush-border ferrireductase) reduces Fe³⁺ → Fe²⁺
2. DMT1 (Divalent Metal Transporter 1) on the apical (luminal) surface transports Fe²⁺ into the enterocyte
3. Inside the enterocyte: iron can be (a) stored temporarily as ferritin (and lost when the enterocyte is shed at the villous tip every 3–5 days) or (b) exported across the basolateral membrane
4. Ferroportin (FPN1 / SLC40A1) exports iron from the basolateral surface into the portal blood — this is the ONLY known iron export protein in the body
5. Hephaestin (a copper-containing ferroxidase) co-localises with ferroportin and oxidises Fe²⁺ → Fe³⁺ for transferrin loading
Factors that enhance non-haem absorption:
- Vitamin C (ascorbic acid): reduces Fe³⁺ → Fe²⁺, keeps iron soluble — this is why prescribing vitamin C with iron tablets is rational
- Gastric acid: provides the acidic environment for Fe³⁺ solubilisation (patients on PPIs have impaired iron absorption — an important iatrogenic cause of iron deficiency)
- Meat factor: unknown mechanism from flesh foods enhances non-haem absorption
Factors that inhibit non-haem absorption:
- Phytates (cereals, legumes): chelate iron in the lumen before absorption — major reason for iron deficiency in vegetarian populations
- Polyphenols (tea, coffee, red wine): form insoluble iron complexes
- Calcium (dairy, calcium supplements): competes with DMT1
- Phosphates, oxalates: form insoluble iron salts
Clinical implication: Advise iron tablet intake 30 minutes before food, with a glass of orange juice (vitamin C), and separated from tea/coffee/dairy by at least 2 hours — this is not pharmacist trivia, it's physiology.
Iron Absorption Pathways in the Duodenum: Haem vs Non-haem
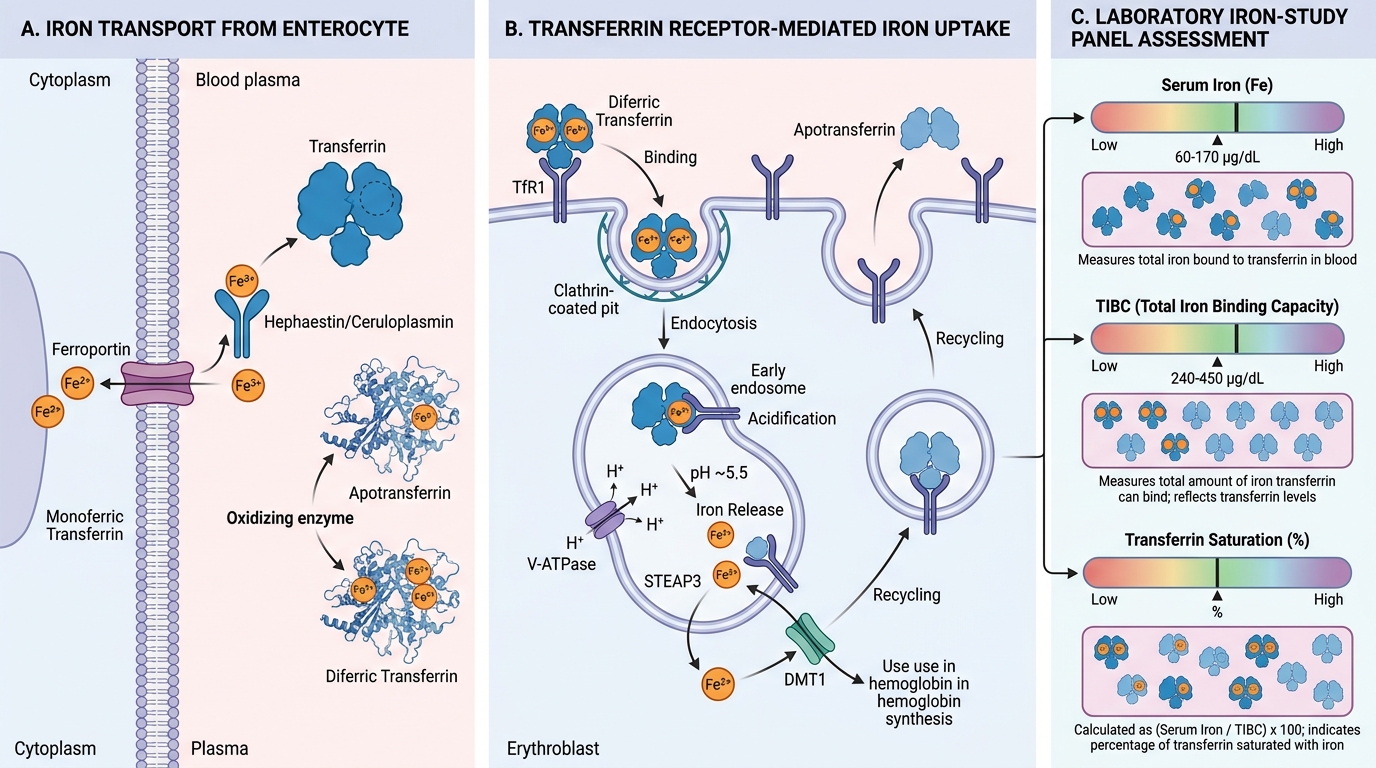
Transport: Transferrin and the Iron-Study Panel
Iron Transport: Transferrin Biology and Laboratory Assessment
Once iron exits the enterocyte via ferroportin, it is immediately bound by transferrin in the plasma. Transferrin is the exclusive plasma iron transport protein.
Transferrin biology:
- Synthesised by the liver (synthesis ↑ when iron stores are low — an inverse relationship with ferritin synthesis)
- Each transferrin molecule has two iron-binding sites → can carry 0, 1, or 2 Fe³⁺ atoms
- Apotransferrin = iron-free transferrin (both sites empty)
- Monoferric transferrin = one site occupied
- Diferric transferrin = both sites occupied (most effective at stimulating erythropoiesis)
Transferrin receptor (TfR1)-mediated uptake:
1. Diferric transferrin binds TfR1 on the cell surface (highest expression on erythroblasts — they need the most iron)
2. Receptor-mediated endocytosis → endosome
3. Endosomal acidification (V-ATPase) causes conformational change → iron released from transferrin → DMT1 transports Fe²⁺ out of endosome into cytoplasm → used for haem synthesis
4. Apotransferrin + TfR1 recycled to cell surface
The iron-study panel — how to read it:
| Parameter | What it actually measures | Normal range |
|---|---|---|
| Serum iron | Fe³⁺ bound to transferrin at time of collection | 60–170 µg/dL |
| TIBC (Total Iron Binding Capacity) | Maximum iron that could bind all plasma transferrin | 240–450 µg/dL |
| Transferrin saturation | Serum iron ÷ TIBC × 100 | 20–50% |
| Serum ferritin | Reflects intracellular ferritin stores (small amount leaks into plasma) | 20–200 ng/mL (women), 20–300 ng/mL (men) |
Key relationship: TIBC is inversely proportional to iron stores. When stores are low, the liver synthesises more transferrin (to try to capture more iron) → TIBC rises. When stores are high (or inflammation is present), transferrin synthesis falls → TIBC falls. This is why TIBC is high in IDA and low in anaemia of chronic disease.
Transferrin saturation interpretation: Normal = ~30%. In IDA: <15%. In iron overload: >70% (risk of free radical generation from non-transferrin-bound iron).
CLINICAL PEARL
The ferritin paradox — your most important laboratory insight in haematology:
Serum ferritin is the best single test for iron-store status — but only in a healthy individual. Ferritin is an acute-phase reactant: any inflammatory state (infection, malignancy, rheumatoid arthritis, inflammatory bowel disease, liver disease, renal failure) causes ferritin to rise independently of iron stores. A ferritin of 80 ng/mL is 'normal' in a healthy person, but it can represent iron-depleted stores in a patient with active Crohn's disease.
Conversely, serum iron is highly diurnal: it peaks in the morning and falls by 30–50% in the evening. It also drops acutely after a meal (non-haem iron absorption creates a transient rise, then storage). This makes serum iron alone nearly useless for diagnosing iron deficiency — always order the full panel, and always specify a fasting morning sample.
Practical rule: In a patient with known chronic inflammation, ferritin <100 ng/mL suggests concurrent iron deficiency; ferritin 100–300 ng/mL is indeterminate; ferritin >300 ng/mL makes iron deficiency unlikely (even if TIBC is not elevated). These thresholds shift clinical decision-making — you may need bone marrow biopsy (Perls' stain) to confirm stores if the picture is mixed.